수면 각성 제어의 수학적 모델링 2편 - 렘수면의 수학
앞선 글에서는 수면과 각성을 오가는 기전에 대해 수학적으로 설명하는 개념적 모델 중 가장 대표적으로 알려져 있는 Two process model과 Sleep-wake flip-flop model을 소개하였다. 이 두 개는 수학적으로 단순화시킨 모델이므로 잠에 들고 깨는 현상을 간단하게 표현해주는 장점이 있지만, 잠든 뒤 수면 단계인 렘수면(REM sleep)과 비렘수면(NREM sleep)에 대한 설명이 불가능한 한계점이 있다.
수면 중 신체는 항상 같은 상태를 유지하는 것이 아니라, 뇌파와 안구움직임 등의 변화를 기준으로 렘수면과 비렘수면을 오가는 주기를 반복한다. 꿈을 꾸는 단계인 렘수면은 취침 후 약 두 시간 후부터 나타나기 시작하고, 수면 후반부로 가면서 그 빈도와 길이가 늘어난다. 반면, 신체의 회복 과정이 일어나는 깊은 수면에 해당하는 비렘수면은 수면 전반부에 더 길고 깊게 나타난다. 수면 주기에 대한 생물학적 현상이나 의학적인 의미는 본 연재물의 취지에 맞지 않으므로 생략하기로 하고, 이 글에서는 수면 주기를 구성하는 렘수면과 비렘수면 사이를 오가는 현상에 대한 수학적인 모델링과 관련된 이야기를 다루어보기로 한다.

꿈을 꾸는 렘수면과 피로를 회복하는 비렘수면 사이를 오가는 현상을 과연 수학적으로 표현할 수 있을까? 우선 어떤 기전으로 수면 중에 렘수면이 시작되고 끝나는지 알아보자.
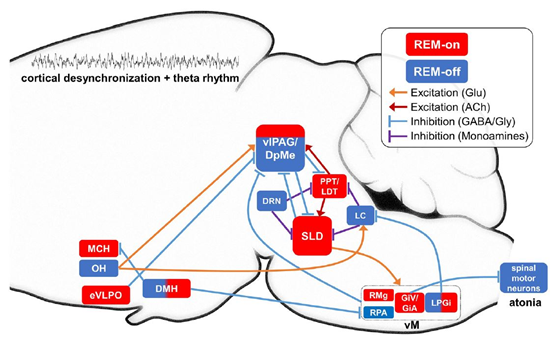
위 그림은 렘수면을 시작하고 종료하는데 작용하는 복잡한 뇌신경 회로를 그린 것이다.[1] 뇌의 여러 부위에서 여러 신경전달물질이 상호작용하면서 렘수면에 영향을 끼치게 된다. 전체적으로 복잡한 신경회로이지만, 이 부분 중 렘수면을 유도하거나 종료시키는데 작용하는 부분만 살펴보면 아래와 같다.[2]
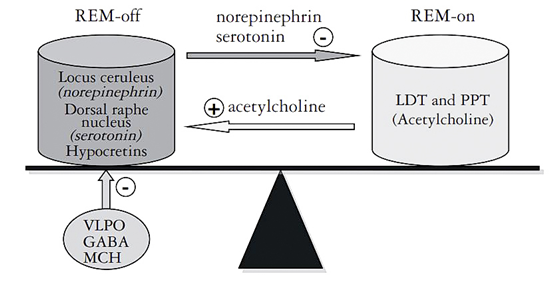
기본적으로 뇌의 laterodorsal tegmental nucleus(LDT)와 pedunculopontine tegmental nucleus(PPT)에서 분비되는 아세틸콜린(acetylcholine)이 렘수면이 발생하도록 유도한다. 반면 locus coeruleus(LC) 와 dorsal raphe nucleus(DR, DRN)에서 분비되는 노르에피네프린(norepinephrine, NE)과 세로토닌(serotonin, 5-HT)등의 모노아민(monoamine) 계열 신경전달물질들이 LDT와 PPT에 억제 작용을 해서 렘수면을 방해한다.
아직도 복잡하다면, 이 신경전달 물질들의 상호작용을 한 번 더 단순화시켜 요약해보자.
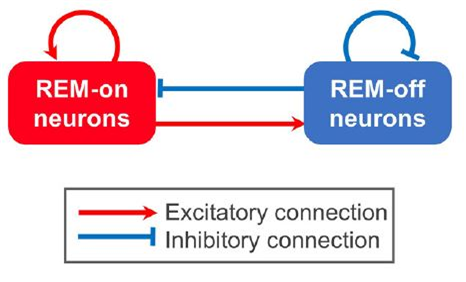
렘수면을 유도하는 REM-on neuron 그룹을 좌측에, 그리고 렘수면을 억제하는 REM-off neuron 그룹을 우측에 그렸다. 언뜻 보면 대칭인 그림 같지만, REM-on neuron에서 나오는 신경전달물질들은 활성화작용(excitatory reaction)만을 유발하고, REM-off neuron에서 분비되는 신경전달물질은 억제작용(inhibitory reaction)을 한다. REM-on neuron이 활성화되면 자신도 스스로 활성화시키지만, REM-off neuron도 동시에 활성화시킨다. 반면 REM-off neuron 은 REM-on neuron을 억제시킨다. [1]
이는 마치 정글 속의 약육강식 관계과도 같다. 무슨 말인가 하면, 먹이(prey)는 생존을 해서 개체수를 늘이려 하는데, 먹이가 늘어나면 포식자(predator)도 덩달아 잘 먹고 개체 수가 늘어가게 된다. 그런데 포식자 개체 수가 많아지면 결국 먹이의 개체 수는 그만큼 줄어들게 된다. 그리고 먹이의 개체 수가 줄어들게 되면 포식자도 덩달아 적어진다. 그리고는 처음처럼 다시 먹이의 개체수가 늘어가는 사이클이 무한 반복되지 않을까? 이 현상에 빗대자면 REM-on neuron은 prey가 되고, REM-off neuron은 predator에 해당한다. 정글의 법칙이 수면주기에도 적용이 된다니 놀랍지 않은가?
더 놀라운 것은 먹이-포식자 관계에 REM-on neuron과 REM-off neuron 관계를 대입함으로써 이 사이 관계를 수학식으로 풀어낼 방법이 생겼다는 점이다. [3][4]

이것은 약육강식 정글의 법칙을 수학적으로 표현한 Lotka-Volterra equation(로트카-볼테라 공식)이다. 이 공식에서 a,b,c,d 는 모두 양수이며, 먹잇감 X는 스스로 증식하고자 하며(aX), 증식할수록 포식자 수가 많아지므로 그에 따라 개체 수가 줄어든다(-bXY). 포식자 Y는 개체 수가 많아질수록 경쟁이 심해지므로 스스로 개체 수가 제한되고(-cY), 먹잇감이 많아질수록 번식한다(+dXY).
렘수면과 관련된 신경전달물질 상호작용을 요약해 보여줬던 그림에 이 공식을 도입해서 이해해보자.
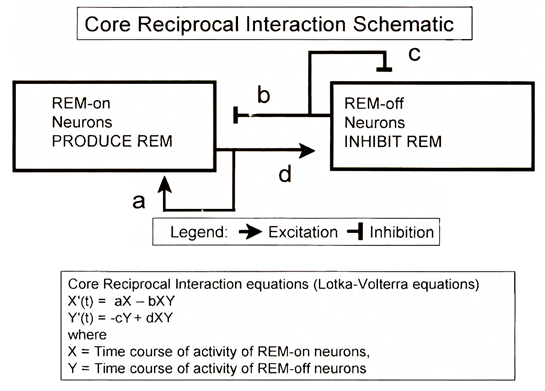
이 모식도를 다시 보면 공식에서 양쪽 신경 그룹이 상호작용하는 a,b,c,d 가 어떤 의미를 갖는지 이해할 수 있다.[5]
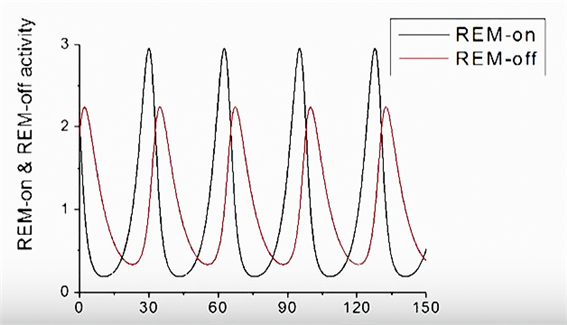
그리고 이 공식대로 그래프를 그려보면, 위와 같이 렘수면유도 신경(먹잇감)과 렘수면억제 신경(포식자)이 주기적으로 증감을 반복하게 된다.[6]
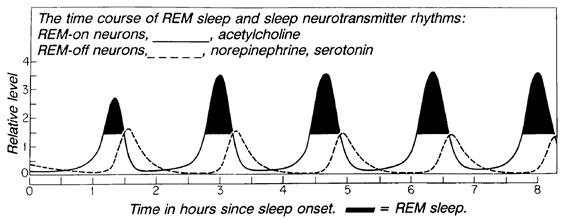
Lotka-Volterra equation으로 REM-on neuron 과 REM-off neuron의 그래프를 그렸을 때 검정색으로 칠한 부분이 실제 렘수면이 일어나고 있는 시간에 해당한다.참고문헌5) 수면 후반부로 갈수록 렘수면의 길이가 조금씩 늘어나는 현상이 실제 수면에서 관찰되는 현상과 일치하는 것을 볼 수 있다.
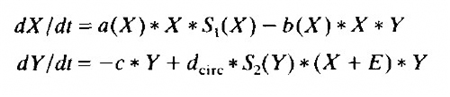
실제 뇌신경계에 작용하는 다른 변수들과, 생체리듬(circadian rhythm)과 관련된 변수를 추가하여 나중에 이 공식은 위와 같이 약간 더 복잡해졌지만(limit cycle equation) 처음의 로트카 볼테르 공식에서 크게 벗어나진 않는다.[6]
1편에서 수면과 각성을 오가는 현상에 대한 수학적 모델을 찾아봤고, 이번 글에서는 수면 중 수면주기에 따라 렘수면과 비렘수면이 번갈아 나타나는 현상을 수학적으로 풀어보았다.
참고문헌
[1]CharlotteHéricé. "Circuit mechanisms and computational model of REM sleep" euroscience Research. 2019
[2]Rosa Hasan. "Updates on the sleep-wake cycle" Sleep Science, 2011
[3] Wipedia, Lotka Volterra equation, Avalible:https://ko.wikipedia.org/wiki/%EB%A1%9C%ED%8A%B8%EC%B9%B4-%EB%B3%BC%ED%85%8C%EB%9D%BC_%EB%B0%A9%EC%A0%95%EC%8B%9D
[4]Department of Methemetical Sciences, Durham University,Lotka Volterra equation, Avalible:https://www.maths.dur.ac.uk/~ktch24/term1Notes(10).pdf
[5]R W McCarley. "Mechanisms and models of REM sleep control" 2004
[6]R.W. McCarley, S.G. Massaquioe, "Neurobiological structure of the revised limit cycle reciprocal interaction model of REM cycle control" Journal of Sleep Research. 1992
그림1:https://www.snorelab.com/the-architecture-of-sleep/
그림2,4:https://www.sciencedirect.com/science/article/pii/S0168010218303833
그림3:http://www.sleepscience.org.br/details/94/en-US/updates-on-the-sleep-wake-cycle
그림5,7,9:https://onlinelibrary.wiley.com/doi/epdf/10.1111/j.1365-2869.1992.tb00026.x
그림6,8:http://www.architalbiol.org/index.php/aib/article/view/142429/376
Cover : www.ted.com/talks/matt_walker_how_sleep_affects_your_emotions#t-1599